A cura di:
Assunta Esposito
Dipartimento di Scienze della Vita, Seconda Università di Napoli
Sandro Strumia
Dipartimento di Scienze Ambientali, Seconda Università di Napoli
Rilevamento Floristico-Sociologico
In una prima fase sono stati esaminati i documenti scientifici e di altra natura disponibili al fine di programmare le indagini sul territorio: l’analisi critica delle immagini aeree e delle informazioni riportate nel formulario del SIC hanno fornito le prime indicazioni sulla possibile localizzazione dei rilievi. Sopralluoghi preliminari in campo sono stati inoltre condotti per validare i risultati delle analisi critiche dei documenti pregressi e per individuare le aree da rilevare. Inoltre sono stati effettuati rilievi preliminari per verificare eventuali dubbi o difficoltà interpretative delle istruzioni relative al riempimento dei campi delle schede di rilevamento fornita dall’Università di Cagliari.
I rilievi floristico-vegetazionali sono stati effettuati in quelle fitocenosi che, sulla base dei rilievi preliminari, si configuravano come potenziali habitat, da confermare grazie proprio ai rilievi floristico-vegetazionali.
I rilievi sono stati fatti utilizzando il classico metodo fitosociologico e compilando le “Schede di rilevamento floristico-sociologico” predisposte dall’Università di Cagliari. I rilievi sono stati distribuiti spazialmente in modo da comprendere nella maniera più esaustiva tutte le fitocenosi che rappresentasserro potenziali habitat; considerato il tipo di rilievo e le caratteristiche del sito, per il campionamento è stato utilizzato un disegno randomizzato a blocchi, dove ogni blocco era rappresentato da una comunità vegetale che per complessità strutturale e composizione floristica costituiva un’unità di campionamento rilevabile. Altri rilievi sono stati condotti in habitat non ascrivibili a quelli dell’All. I della Direttiva CEE 92/43, ma ritenuti interessanti in quanto comunque in contatto catenale in aree a ridosso della pineta con vegetazione dominata da terofite.
Per permettere l’ interpolazione dei dati floristico-vegetazionali con quelli di altra natura, parte dei rilievi sono stati eseguiti, dove possibile, in corrispondenza dei rilievi per le schede “rilevamento – taxon”; in particolare, alcuni rilievi sono stati effettuati per caratterizzare i popolamenti in cui risultava presente Daphne sericea Vahl, entità ritenuta interessante dal punto di vista conservazionistico in quanto citata nella Lista Rossa per la Campania anche se col valore (LR) (Conti et al., 1997).
Tutti i rilievi sono stati georeferenziati utilizzando un GPS Garmin E-trex H.
Le piante non identificabili in campo sono state raccolte e determinate in laboratorio con l’ausilio di uno stereomicroscopio Leica MZ 12,5 ed utilizzando le chiavi dicotomiche di Pignatti (1982) e Tutin et al (1964-80; 1993).
Tutti i campioni raccolti sono stati essiccati e sono attualmente conservati presso l’erbario della Seconda Università di Napoli (CASERTA). I dati raccolti sono stati informatizzati utilizzando lo stesso foglio di lavoro Excel utilizzato per il rilevamento in campo.
Per effettuare tutte le elaborazioni successive, i dati floristici dei rilievi della vegetazione sono stati inseriti anche nel geodatabase relazionale sviluppato presso il Laboratorio di Botanica Sistematica ed Ambientale del Dipartimento di Scienze Ambientali della Seconda Università degli Studi di Napoli (Santangelo et al., 2008). In questa banca dati floristica risultano disponibili i dati tassonomici relativi a tutte le entità della flora campana secondo quanto proposto nella Check-list della Flora Vascolare Italiana (Conti et al., 2005; 2007) ed i relativi dati distributivi (Santangelo, 2005; Conti et al., 2007); sono inoltre riportate le informazioni relative al loro eventuale interesse conservazionistico, in accordo con Conti et al. (1997), e con le normativo sia a livello internazionale (inclusione negli allegati alla convenzione di Berna e/o alla direttiva CEE 92/43) che locale (legge regionale 40/94). Nel database sono infine disponibili le informazioni di tipo corologico ed ecologico (Pignatti, 1982), le fonti bibliografiche relative ai dati distributivi sulla flora della regione Campania pubblicate dal 1950 in poi (Scoppola e Magrini, 2005) e più antiche (Del Guacchio e Gargiulo, 2006). Il database così strutturato permette una veloce verifica dell’interesse scientifico e protezionistico di elenchi floristici ed una veloce realizzazione di elaborazioni di vario tipo, tra cui anche matrici di dati multivariati.
I dati informatizzati sono stati sottoposti ad analisi statistica utilizzando tecniche di analisi multivariata utilizzando il software SYN-TAX 2000 (Podani, 2001). Per la classificazione è stato utilizzato l’indice di Jaccard su base qualitativa come indice di dissimilarità ed il legame medio (UPGMA) come criterio agglomerativo. Le informazioni su caratteristiche ambientali, struttura della comunità vegetale, forme biologiche, corotipi e le informazioni sull’eventuale grado di protezione delle singole specie sono state utilizzate come strumento interpretativo dei diagrammi ottenuti.
I dati raccolti sono stati analizzati per caratterizzare in termini sintassonomici le fitocenosi rilevate. I risultati sono stati infine confrontati con le descrizioni degli habitat riportati nell’ “Interpretation Manual of European Union Habitats – EUR 27” del luglio 2007 e nelle schede del “Manuale Italiano di interpretazione degli habitat della Direttiva 92/43/CEE” (disponibile sul sito ufficiale del Ministero dell’Ambiente e della Tutela del Territorio e del Mare), per confermare da una parte la presenza degli habitat già segnalati ed eventualmente riconoscere e segnalare la presenza di nuovi habitat.
Rilevamento taxon
Questa attività ha come obiettivo lo studio fenologico/demografico di specie strutturali degli habitat selezionati e che rivestano particolare importanza come specie endemiche e/o di interesse fitogeografico a livello locale. Ciò allo scopo di poterne valutare lo stato di conservazione ed identificare con maggiore dettaglio quantitativo e qualitativo le cause di degrado degli habitat e pianificare con maggiore precisione le azioni di conservazione in situ necessarie. Le analisi fenologiche e demografiche sono finalizzate ad individuare i trend demografici delle singole popolazioni, in particolare i tassi di natalità, mortalità e recruitment oltre alla distribuzione spaziale del popolamento. Il corretto funzionamento di tutto il ciclo biologico è fondamentale per garantire la riproduzione sessuale e, quindi, la conservazione della specie nel tempo. E’ sufficiente, infatti, che una sola delle fasi (fioritura, impollinazione, fecondazione, disseminazione, germinazione) venga in qualche modo alterata per avere una ripercussione negativa sull’efficienza della riproduzione e quindi sulla possibilità di conservazione di una determinata specie.
A tale riguardo i rilievi che possano fornire informazioni utili ed esaustive devono essere condotti su un intervallo temporale che copra almeno un intero ciclo vitale della specie oggetto di interesse. Tuttavia è indispensabile condurre indagini su più cicli consecutivi al fine di poter meglio valutare e discriminare se le eventuali cause di alterazione delle varie fasi del ciclo vitale di una specie siano da attribuire a fattori climatici e/o antropici
Per queste ragioni e tenuto conto dei tempi limitati per condurre tali indagini si è fatto riferimento ai dati floristici parziali disponibili al momento di avvio della ricerca al fine di selezionare le specie più rappresentative e che rispondessero ai requisiti di interesse conservazionistico così come previsto dal progetto.
Lo studio ha previsto, quindi, una prima fase di analisi delle informazioni raccolte da questo gruppo di lavoro in una ricerca avviata di recente (Croce et al. 2009).. Per quanto riguarda le specie caratteristiche degli habitat lo studio è stato condotto su: Juniperus oxycedrus L. macrocarpa (Sibth. & Sm.) Neilr. e Crucianella maritma L. Per quanto riguarda le specie di interesse fitogeografico l’analisi è stata condotta su Daphne sericea Vahl, specie arbustiva ritenuta interessante dal punto di vista conservazionistico in quanto citata nella Lista Rossa per la Campania anche se col valore (LR) (Conti et al., 1997).
Per il rilevamento della Daphne sericea l’analsi del taxon e dello stato fenologico è stata condotta, ogni mese, su tutti gli individui presenti nel SIC e la cui posizione è stata georeferenziata utilizzando un GPS Garmin iQUE 3600 consentendo, in questo modo, di individuare con precisioni ogni individuo nei successivi campionamenti e di condurre un’analisi della distribuzione spaziale del popolamento e dei fattori ambientali stazionali al fine di individuare eventuali fattori critici.
Parallelamente ai rilievi in campo è stata condotta una ricerca bibliografica finalizzata alla individuazione di eventuali dati di letteratura relativi alla distribuzione, biologia ed ecologia di Daphne sericea.
Per quanto riguarda Crucianella maritima L. si è proceduto ad individuare e georeferenziare le poche stazioni presenti nel SIC e costitite da singoli indiviui o gruppi di poche unità utilizzando un GPS Garmin iQUE 3600 per consentire anche in questo caso un’analisi spaziale del popolamento estremamente rarefatto e discontinuo. Le osservazioni fenologiche sono state condotte su un campione di 10 individui selezionati a random nell’ambito delle piccole stazioni rilevate lungo tutta l’area del SIC.
Per quanto riguarda Juniperus oxycedrus L. macrocarpa (Sibth. & Sm.) Neilr. sono stati effettuati rilievi del taxon e della fenologia all’interno di tre aree di rilievo permanenti di 50 m x 50m selezionate nell’ambito dell’habitat occupato dal ginepreto come di seguito indicato: 1) nella parte più a nord prossima alla foce del Garigliano; 2) nella zona centrale in prossimità del villaggio internazionale; 3) nella parte più a sud in prossimità del villaggio svedese.
Risultati
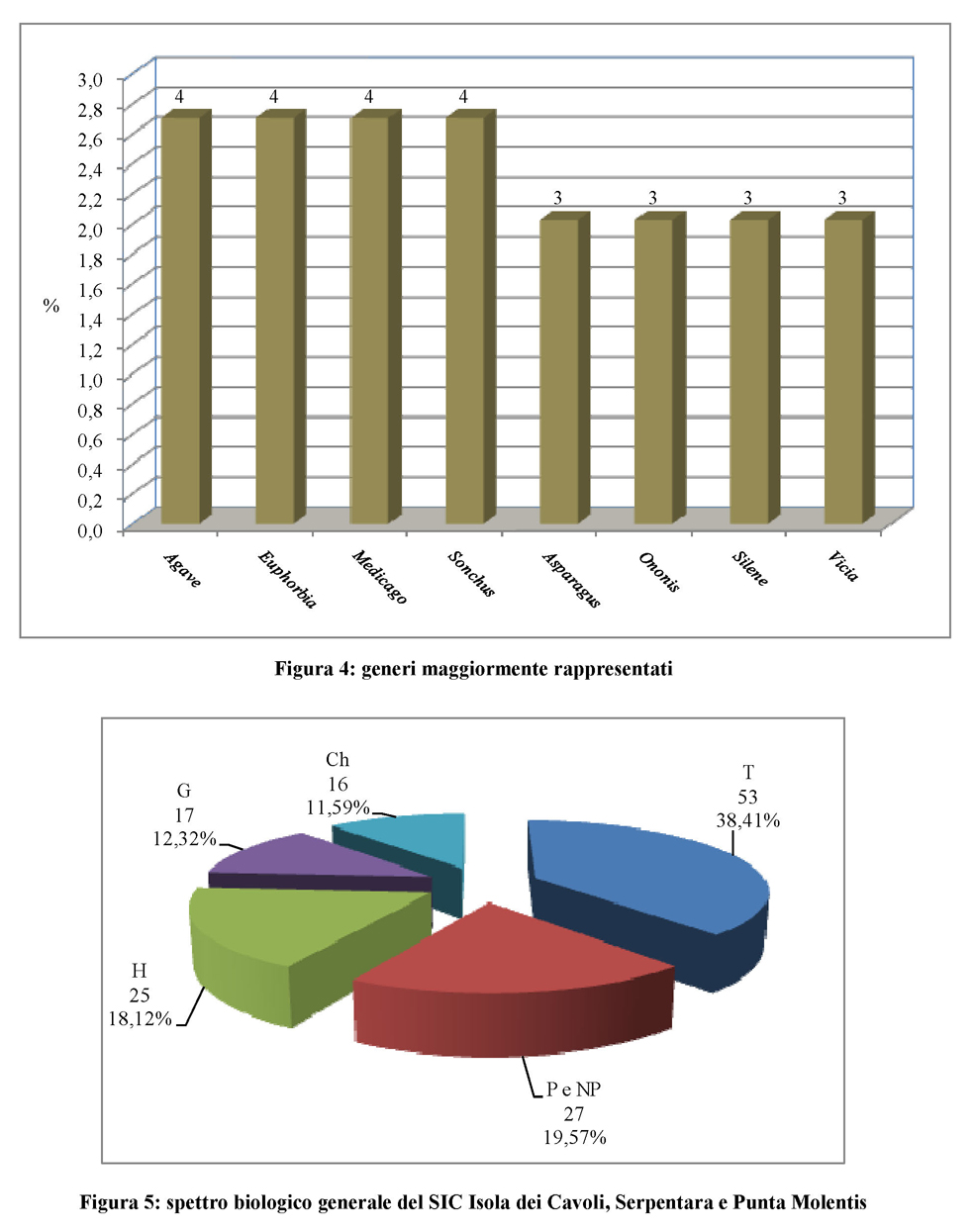
Grazie all’attività di rilevamento in campo sono stati effettuati 83 rilievi floristico-sociologici. L’informatizzazione dei dati dei rilievi ha generato una matrice di 83 rilievi x 187 specie. A queste 187 specie sono state aggiunte altre 68 entità ritrovate nel sito al di fuori degli habitat indagati o nell’ambito delle ricerche svolte in precedenza da questo gruppo di lavoro.
Elenco floristico
In totale sono state rilevate 255 entità. Una verifica dell’entità ritrovate permette di affermare che nessuna delle entità rilevate è compresa nell’Allegato II della Direttiva CEE 92/43.
Nelle tabelle 4 e 5 sono riportati rispettivamente gli spettri biologico e corologico.

Lo spettro biologico evidenzia il carattere di forte mediterraneità del sito con una elevata percentuale di Terofite (42.3%); ben rappresentata anche la componente di Emicriptofite (22.0%) il cui significato potrebbe essere legato più all’uso del territorio piuttosto che a motivi legati al clima. L’analisi dello spettro corologico fornisce una conferma a questa prima interpretazione dei dati: la componente Mediterrannea s.l. (Steno- ed Euri-Medit.) supera il 50% del totale delle entità rinvenute, confermando la coerenza della componente floristica con le caratteristiche climatiche dell’area. Nello stesso tempo però è opportuno sottolineare che la terza tipologia di corotipo in ordine di frequenza percentuale è rappresentato dalle specie ad Ampia Distribuzione (22.7%) evidenziando la profonda antropizzazione dell’area. Infatti tra queste si registrano molte entità tipiche di aree ruderali o infestanti delle colture come Chenopodium album L. subsp. album, Portulaca oleracea L. s.l., Fumaria officinalis L. subsp. officinalis, Aphanes arvensis L., Tribulus terrestris L., Euphorbia helioscopia L. subsp. helioscopia, Daucus carota L. subsp. carota, Convolvulus arvensis; numerose anche le specie esotiche coltivate come Eucalyptus camaldulensis Dehnh. o Acacia retinoides Schltdl., quest’ultima anche con caratteristiche di invasività e spontaneizzazione evidente. Particolare rilevanza assume la presenza di Carpobrotus acinaciformis (L.) L. Bolus, riconosciuta come estremamente pericolosa per la conservazione ed il mantenimento delle comunità vegetali delle spiagge sabbiose, in quanto capace di formare densi tappeti molto resistenti, a scapito delle specie autoctone.
Nell’ area d’indagine sono state rilevate anche 7 entità endemiche; si tratta di:
- Dianthus balbisii Ser. subsp. liburnicus (Bartl.) Pignatti
- Dianthus carthusianorum L. subsp. tenorei (Lacaita) Pignatti
- Biscutella maritima Ten.
- Senecio gibbosus (Guss.) DC. subsp. bicolor (Willd.) Peruzzi, N.G. Passal. & Soldano
- Phleum hirsutum Honck. subsp. ambiguum (Ten.) Tzvelev
- Ophrys exaltata Ten. subsp. tyrrhena (Gölz & H.R. Reinhard) Del Prete
- Ophrys crabronifera Mauri
Due entità presenti nell’elenco risultano inoltre inserite nella lista rossa regionale con i seguenti valori di vulnerabilità:
- Daphne sericea Vahl (LR)
- Dianthus balbisii Ser. subsp. liburnicus (Bartl.) Pignatti (LR)
Relativamente al valore protezionistico bisogna inoltre sottolineare che oltre alle due entità citate, nell’area indagata sono state ritrovate altre 14 entità appartenenti alla famiglia delle Orchidacee e come tali incluse nell’Allegato della L.R. 40/94 e sottoposte a vincolo di tutela; altre specie rinvenute nel corso della ricerca che compaiono nello stesso allegato sono Typha latifolia L. e Laurus nobilis L., anche se per quest’ultima specie andrebbe verificato il reale significato della sua presenza.
Rilevamento taxon
Daphne sericea Vahl (Fig. 2), appartente alla famiglia delle Thymeleaceae, è specie rara in quanto presente in poche località della regione mediterranea orientale ed in Italia a Marittimo, nel Gargano, in Abruzzo presso Popoli e Sulmona, sulle Ponziane ed in alcune zone della macchia tirrenica, dalla maremma toscana al napoletano (Pignatti, 1982). Le popolazioni residuali della Campania sono attualmente segnalate solo nel SIC Pineta di Foce Garigliano e all’interno della Riserva Naturale di Castelvolturno (CE) ( Croce et al. 2009; Esposito et al. 2009).
Studi recenti sulla biologia fiorale ed ecologia di popolazioni campane e toscane (Aronne, e Wilcock 1996; Martinez-Pallè, E. 1998; Barbi, 2008). hanno evidenziato che il ciclo riproduttivo di D. sericea comincia all’inizio del mese di marzo, con lo sviluppo delle gemme fiorali, e si conclude nel mese di giugno, con la maturazione dei frutti. I fiori, raggruppati in fascetti apicali, sono monoclini con caratteristica colorazione rosa brillante e molto profumati per essere fortemente attrattivi per molti tipi di insetti sia diurni che notturni (Aronne, e Wilcock 1996). Il frutto si sviluppa velocemente fino ad avere, a maturazione avvenuta, una colorazione arancione intenso ed essere disperso sia da uccelli che formiche (Barbi, 2008). Questi studi hanno evidenziato nel ciclo riproduttivo di D. sericea fasi critiche rappresentate dalla scarsa produzione di frutti maturi, dalla germinazione dei semi e dal successivo attecchimento delle plantule.

Nel corso dello studio da noi condotto, l’analsi di rilevamento in campo ha portato all’individuazione, su tutta l’area occupata dall’habitat 2250* – Dune costiere con Juniperus spp., di 45 individui adulti.
Tale elaborazione mostra in modo evidente una chiara distribuzione a macchia (patchiness) a tratti puntiforme alternati a piccoli nuclei di una decina di individui o a settori con completa assenza di individui. Questa distribuzione spaziale fortemente discontinua è certamente attribuibile, in primo luogo, a cause antropiche legate principalmente, in alcuni settori, al forte rimaneggiamento, ai fini dell’insediamento di lidi balneari, dell’habitat idoneo a garantire la presenza di tale specie
Viceversa l’estrema rarefazione del popolamento potrebbe essere attribuita alle mutate condizioni ecologiche idonee alla svolgimento dell’intero ciclo vitale della specie. I dati di letteratura citati in precedenza hanno evidenziato una criticità nella fase di germinazione e attecchimento delle plantule da associare con molta probabilità ad un progressivo inaridimento delle condizioni ambientali. Del resto già Tenore nel 1830 rese evidente il fatto di ritrovare questa specie sempre associata alla presenza di acqua ed in particolare presso le sponde dei ruscelli e nelle zone paludose. Considerata la vasta opera di bonifica dei litorali paludosi avvenuta attorno agli anni ’50, si può ipotizzare che la distribuzione di questa specie, già limitata all’inizio del XX secolo, si sia ulteriormente e fortemente ridotta a causa delle opere di drenaggio dei suoli e, negli ultimi decenni, amplificata a causa degli insediamenti turistici lungo la fascia litoranea.
La delicatezza di questi ambienti fa sì che una loro frammentazione anche parziale e localizzata, porti al progressivo degrado dell’intero ecosistemi. In particolare tagli nel cordone l’apertura di passaggi verso la spiaggia, fanno sì che i venti salmastri e le grandi masse di sabbie sciolte, non più frenati dalle strutture sabbiose consolidate della prima duna, si spostino nell’entroterra minando l’integrità della vegetazione retrostante e nel caso specifico dell’habitat in cui è presente Daphne sericea.
Il delicato equilibrio è inoltre confermato, anche nel nostro caso, dall’aver rilevato su tutta l’area indagata la completa assenza di individui giovani ed in un solo caso la presenza di una plantula.
Per quanto riguarda la fenologia sono stati raccolti dati da ottobre 2009 ad agosto 2010 che hanno permesso di analzzare gli andamenti mensili sulle caratteristiche della biologia come riportato nei grafici seguenti:
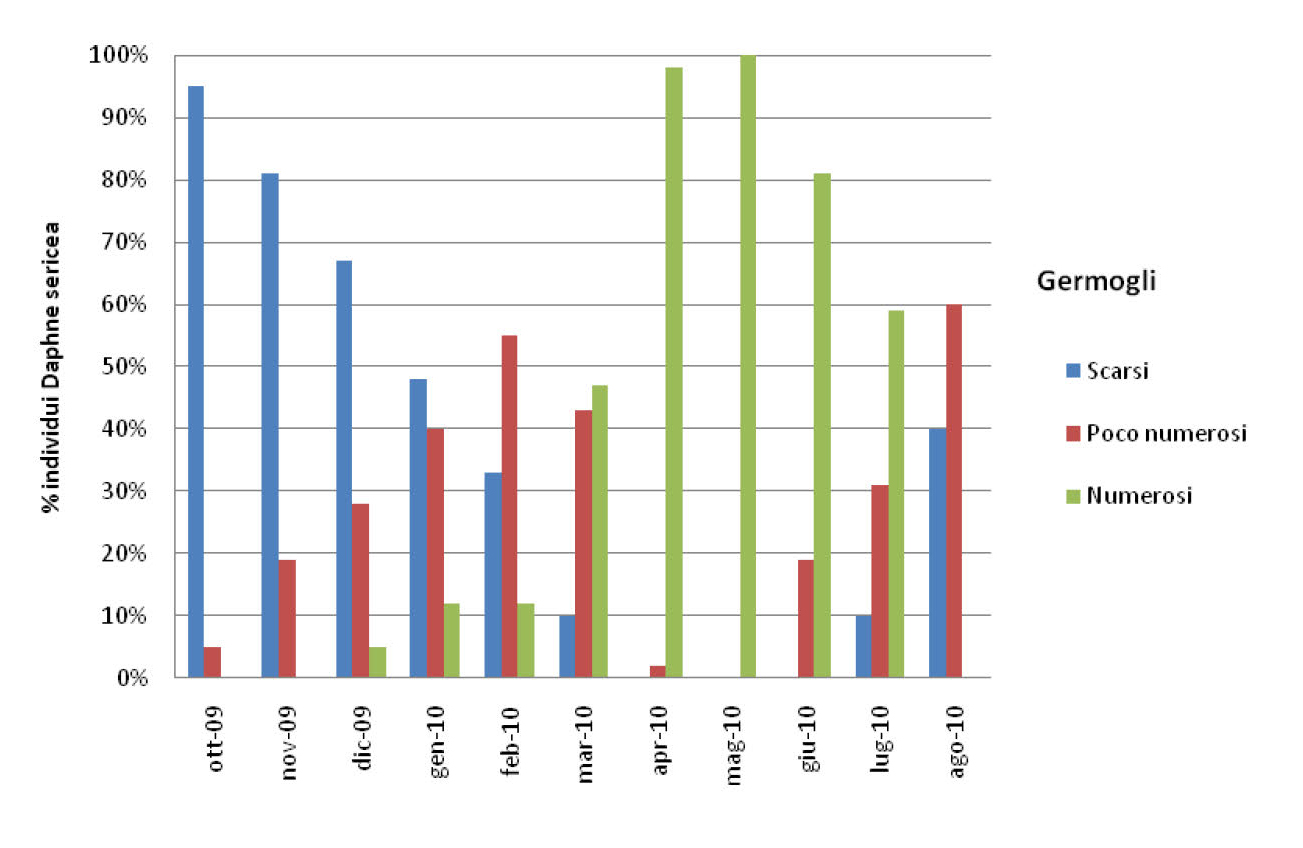

I dati evidenziano che la fase più critica del ciclo vitale è rappresentata dalla fruttificazione. E’ molto probabile, cosi come già evidenziato dagli studi condotti da Scippa 2008, che la particolare distribuzione del popolamento possa indurre gli impollinatori a visitare piante della stessa popolazione diminuendo così il flusso genico inter-popolazione e la variabilità genetica intra-popolazione. Ciò unitamente alle ridotte dimensioni della popolazione costituisce una minaccia evidente della possibilità di sopravvivenza di tale specie.
In tale ottica azioni mirate alla salvaguardia e conservazione di tale entità si configurano come obiettivi prioritari da porre in essere in tale progetto. Tra esse le misure di intervento ipotizzabili prevedono: il prosieguo delle attività di rilevamento della fenologia della specie ed in particolare del germoplasma; l’allestimento di un vivaio per la coltivazione ex situ e la produzione di piantine da utilizzare in una fase di arricchimento del popolamento soprattutto nelle aree più povere; l’utilizzo del materiale coltivato anche a scopo ornamentale dato l’idoneo portamento della specie. Per quanto riguarda Crucianella marittima la georeferenziazione dei rilievi evidenzia una distribuzione continua su tutta l’area sebbene le stazioni risultino fortemente compromese e costituite da individui presenti a piccoli gruppi o in aluni casi isolati. Per quanto riguarda la fenologia sono stati raccolti dati da gennaio 2010 ad agosto 2010 che hanno permesso di analizzare gli andamenti mensili sulle caratteristiche della biologia fiorale come di seguito riportato: 
I dati evidenziano una discreta presenza di fioritura caratterizzata però da una scarsa produzione di fiori e successivamente di frutti maturi. Va sottolineato tuttavia che tali dati non sono esaustivi poiché relativi ad un solo ciclo annuale e quindi espressione delle caratteristiche climatiche osservate nell’anno ri rilevamento.
Per quanto riguarda Juniperus oxycedrus L. macrocarpa (Sibth. & Sm.) Neilr. Il rilevamento del taxon condotto da ottobre 2009 ad agosto 2010 ha evidenziato che nelle aree di rilievo permanente la presenza di giovani plantule si rileva in numero piuttosto limitato nel periodo invernale primaverile ma con una successiva mortalità nel periodo estivo. Per quanto riguarda la fenologia sono stati raccolti dati da ottobre 2009 ad agosto 2010 che hanno permesso di analizzare gli andamenti mensili come di seguito riportato. 

I dati fenologici non evidenziano criticità nella produzione di fiori e successivamente di frutti maturi . Anche in questo caso va sottolineato tuttavia che tali dati non sono esaustivi poiché relativi ad un solo ciclo annuale e quindi espressione delle caratteristiche climatiche osservate nell’anno ri rilevamento.
Conclusioni
Le analisi hanno permesso di caratterizzare le fitocenosi indagate e di evidenziarne lo stato di salute; inoltre alcuni sono emersi alcuni aspetti estremamente interessanti ai fini gestionali:
- La componente floristica appare alterata da un elevato numero di specie ad Ampia distribuzione (22,7 % del totale) che testimoniano il forte grado di antropizzazione dell’area. Tra queste specie numerose sono quelle ruderali ed infestanti che contribuiscono ad abbassare la naturalità dell’area; ai fini gestionali preme sottolineare la presenza di alcune avventizie come Carpobrotus acinaciformis (L.) L. Bolus, le cui proprietà invasive possono contribuire ad alterare profondamente le comunità psammofile autoctone, mettendo in pericolo la sopravvivenza degli habitat a dominanza di specie erbacee. La presenza e la distribuzione spaziale di questa specie andrebbe monitorata nel tempo per verificarne la diffusione nell’area. Questa come altre specie coltivate dovrebbero lentamente essere eliminate per aumentare la naturalità dell’area.
- Nell’area sono state rinvenute 7 entità endemiche o subendemiche a testimoniare il significato dell’area in termini di conservazione della biodiversità; due entità (Daphne sericea Vahl e Dianthus balbisii Ser. subsp. liburnicus (Bartl.) Pignatti risultano inoltre inserite nella lista rossa regionale, entrambe con valore di vulnerabilità (LR).
- la vegetazione psammofila più prossima al mare (ascrivibile agli habitat: 2110 – Dune mobili embrionali; 1210 – Vegetazione annua delle linee di deposito marine) ma anche quella che dovrebbe occupare le aree prossime alle dune più stabilizzate (2210 – Dune fisse del litorale (Crucianellion; 2230 Dune con prati dei Malcolmietalia), è ridotta a piccoli lembi ed appare fortemente alterata sia in termini di composizione floristica che strutturale, al punto che in alcuni casi se ne riconosce più la potenzialità che la reale presenza.
La ricerca ha comunque permesso di riconoscere un habitat (2230 Dune con prati dei Malcolmietalia), non precedentemente segnalato nel formulario del SIC.
- L’ habitat 2270* – Dune con foreste di Pinus pinea e/o Pinus pinaster possiede la copertura percentuale più elevata. Malgrado si tratti chiaramente di un rimboschimento, la sua stessa estensione impone alcune considerazioni di carattere gestionale. Anche se sulla base di quanto indicato nel Manuale Italiano, la gestione di questo habitat è piuttosto controversa, si ritiene che nel caso in questione andrebbe cercato un giusto equilibrio tra la conservazione di questo habitat e la sua evoluzione verso altri tipi di habitat. In quest’ottica sarebbe necessario redigere un piano di gestione forestale che da un lato garantisca la sopravvivenza del soprassuolo forestale, ma dall’altro programmi la riduzione della densità degli individui attraverso tagli controllati che favoriscano il recupero della vegetazione autoctona del sottobosco. Altro elemento da considerare è l’attuale assenza di qualsiasi forma programmata di manutenzione del bosco che ha determinato l’accumulo di notevoli quantità di lettiera e di necromassa di varia dimensione e spessore. Questo materiale morto rappresenta un potenziale combustibile, che aumenta il rischio di incendi incontrollabili e particolarmente distruttivi; la rimozione, anche attraverso l’uso di tecniche di fuoco prescritto, della necromassa dovrebbe rappresentare una pratica gestionale utilizzata con regolarità ai fini della salvaguardia dell’intero habitat e di quelli limitrofi.
- Le comunità presentano una forte mosaicizzazione al punto che spesso elementi floristici tipici di un’associazione vegetale – habitat si rinvengono ,col significato di specie ingressive, all’interno dei rilievi effettuati.
- Questa frammentazione del pattern orizzontale determina spesso la difficile attribuzione di una comunità ad un’habitat in quanto oltre ad essere presenti le specie caratteristiche, se ne rinvengono altre che talvolta testimoniano una diversa facies evolutiva dell’habitat (sia in termini degradativi che aggradativi), oppure la possibile presenza di un altro habitat in contatto catenale.
- I motivi delle alterazioni osservate possono essere ricondotti, fondamentalmente, a processi di erosione e calpestio diffusi in tutta l’area; queste azioni contribuiscono a realizzare un effetto isola, dove la normale seriazione della vegetazione anzicchè seguire la direzione perpendicolare alla linea di costa (condizione naturale in assenza di disturbi), si presenta più con una disposizione “radiale” rispetto a queste “isole” più o meno grandi di vegetazione in cui le porzioni centrali sono occupate da vegetazione a dominanza di specie legnose ed ai bordi si incontrano fitocenosi sempre meno strutturate ed ospitanti elementi floristici tipici delle fitocenosi pioniere delle sabbie.
- In tutta l’area risulta evidente l’impatto antropico dovuto presumibilmente alle attività gestionali connesse con il turismo di tipo balneare. Queste attività comprendono sia quelle che insistono sull’area nel momento dell’uso a fini balneari (calpestio da parte dei turisti con evidente conferma o allargamento delle aree di discontinuità tra comunità vegetali, occupazione con strutture più o meno fisse delle aree di pertinenza della vegetazione delle dune), ma anche e soprattutto quelle di tipo preparatorio. Queste ultime infatti spesso determinano effetti non osservabili immediatamente, ma a lunga scadenza e proprio per questo talvolta difficilmente recuperabili; a titolo di semplice esempio si vuole ricordare la pratica diffusa della “pulitura” delle spiagge (in particolare della fascia più prossima al mare in quanto di maggiore interesse ai fini della balneazione) attraverso mezzi meccanici che alterano non solamente la vegetazione esistente, ma anche quella futura attraverso la rimozione di quelle unità di dispersione (frutti, semi, propaguli) essenziali per la sopravvivenza delle specie annuali, ma anche perenni. Spesso tutto ciò viene compiuto dai gestori delle attività turistiche in assoluta buona fede in quanto essi stessi non sono consapevoli dell’ecologia degli ecosistemi che essi sono chiamati a gestire e pertanto dei danni (a volte irreparabili) che causano con pratiche errate. In questo senso sarebbe assolutamente auspicabile prevedere tra le azioni volte alla conservazione degli habitat il coinvolgimento dei gestori di queste attività attraverso corsi di informazione – formazione che li rendano pertecipi del processo di conservazione non solo degli habitat, ma anche di tutto il sistema dunale che per loro rappresenta una risorsa economica prioritaria.
- L’analisi del taxon Daphne sericea ha portato all’individuazione, su tutta l’area occupata dall’habitat 2250* – Dune costiere con Juniperus spp., di 42 individui adulti e di assenza di individui giovani e plantule.
- Al numero piuttosto esiguo di individui che costituiscono il popolamento di Daphne sericea si associa una distribuzione spaziale a macchia (patchiness) con tratti a presenza puntiforme alternati a piccoli nuclei di una decina di individui o a settori con completa assenza di individui.
- la distribuzione spaziale fortemente discontinua di Daphne sericea è certamente attribuibile, in primo luogo, a cause antropiche legate principalmente, in alcuni settori, al forte rimaneggiamento, ai fini dell’insediamento di lidi balneari, dell’habitat idoneo a garantire la presenza di tale specie
- Viceversa l’estrema rarefazione del popolamento potrebbe essere attribuita alle mutate condizioni ecologiche idonee alla svolgimento dell’intero ciclo vitale della specie ed in particolare ad un progressivo inaridimento delle condizioni ambientali.
- La frammentazione anche parziale e localizzata dell’habitat, in particolare tagli l’apertura di passaggi verso la spiaggia, possono costituire una seria minaccia all’integrità della vegetazione retrostante e nel caso specifico dell’habitat in cui è presente Daphne sericea.
- La scarsa rappresenatività e forte compromissione del popolamento a Crucianella marittima che evidenzia una bassa produzione di frutti maturi e l’assenza di plantule.
- L’assenza, almeno per il periodo di rilevamento indagato, della rinnovazione dei popolamenti ad Juniperus oxycedrus L. macrocarpa (Sibth. & Sm.) Neilr evidenzia anche in questo caso una forte alterazione dell’habitat imputabile in parte alla forte erosione del litorale che espone questi popolamenti a condizioni edafiche critiche.
- Azioni mirate alla salvaguardia e conservazione di tale entità si configurano come obiettivi prioritari da porre in essere in tale progetto. Tra esse le misure di intervento ipotizzabili prevedono: il prosieguo delle attività di rilevamento della fenologia della specie ed in particolare del germoplasma; l’allestimento di un vivaio per la coltivazione ex situ e la produzione di piantine da utilizzare in una fase di arricchimento del popolamento soprattutto nelle aree più povere; l’utilizzo del materiale coltivato anche a scopo ornamentale dato l’idoneo portamento della specie.
Bibliografia di riferimento
Croce A., Strumia S. & Esposito A., 2009. Contribution to the floristic and vegetation knowledge of Garigliano estuary area (CE, southern Italy). Bacchetta G. (ed.) – Biodiversity hotspots in Mediterranean Area. Book of abstract 45° International Congress of SISV & FIP: p. 274.
Del Guacchio E., 2010. Appunti di floristica campana: novità e precisazioni. Informatore Botanico Italiano 42(1): 35-46.
Del Guacchio E., Gargiulo S., 2006. Bibliografia geobotanica della Campania dal XVI secolo fino al 1949. Delpinoa, n.s., 46: 29-50. (2004)
Esposito A., Filesi L., Zumbolo A., Strumia S. 2009. Characterization of vegetation landscape of Castel Volturno Nature Reserve (Campania region, Southern Italy). Bacchetta G. (ed.) – Biodiversity hotspots in Mediterranean Area. Book of abstract 45° International Congress of SISV & FIP: p. .
European Commission DG Environment – Nature and Biodiversity, 2007. Interpretation Manual of European Union Habitats, EUR 27
Martinez-Pallè, E. 1998. Autoecologia e conservazione di Daphne sericea Vahl (Thymeleaceae). Atti del 93° Congresso della Società Botanica Italiana, 1-3 ottobre, p. 96.
Pignatti S., 1982 – Flora d’Italia. 1-3. Edagricole, Bologna.
Podani J. 2001. SYN-TAX 2000. Computer Programs for Data Analysis in Ecology and Systematics. User’s manual. Scientia Publishing, Budapest, Hungary.
Santangelo A., 2005. Dati regionali per la Campania. In: Conti F., Abbate G., Alessandrini A., Blasi C. (eds.), An annotated checklist of the italian vascular flora – F.lli Palombi editori, Roma
Santangelo A., Bronzo E., Croce A., Salvati C., Strumia S., 2008 – Basi di dati per le ricerche floristiche: un esempio per il Matese campano. Inform. Bot. Ital., 40 (1): 59-71.
Scoppola A., Magrini S., 2005. Bibliografia floristica italiana per gli anni 1950-2005. In: Scoppola A., Blasi C. (Ed.), Stato delle conoscenze sulla Flora Vascolare d’Italia- F.lli Palombi editori, Roma.
Tutin T.G., Burges N.A., Chater A.O., Edmondson J.R., Heywood V.H., Moore T.M., Valentine D.H.,Walters S.M. & Webb D.A. (eds.), 1993 – Flora Europaea. 1, 2a Ediz. Cambridge University Press.
Tutin T.G., Heywood V.H., Burges N.A., Valentine D.H., Walters S.M. & Webb D.A. (eds.), 1964-80 -Flora Europaea. 1-5. Cambridge University Press.